Opioids
This is the most heavily abused drug class on record.
Opioids drove 68% of all US overdose deaths in 2024 - roughly 54,000 people, more than any other drug class (CDC, 2025; Overdose Lifeline, 2024). Fentanyl and other synthetic opioids accounted for ~88% of opioid-involved deaths (i.e., roughly 60% of all overdose deaths). Note: Module 10.4 states that fentanyl accounts for ~70% of all overdose deaths using a different DEA dataset and denominator - both figures are in the right range; the exact percentage depends on the data source and year. Per Module 10b's lifetime-dependence data, of people who ever try heroin, about 23% develop dependence - among the highest of any drug class.
Step 1: What Counts as an Opioid
An opioid is any drug that binds one or more of the brain's three classical opioid receptors. Which receptor a particular opioid hits, and how it hits it, determines whether it produces euphoria, dysphoria, analgesia, or something else.
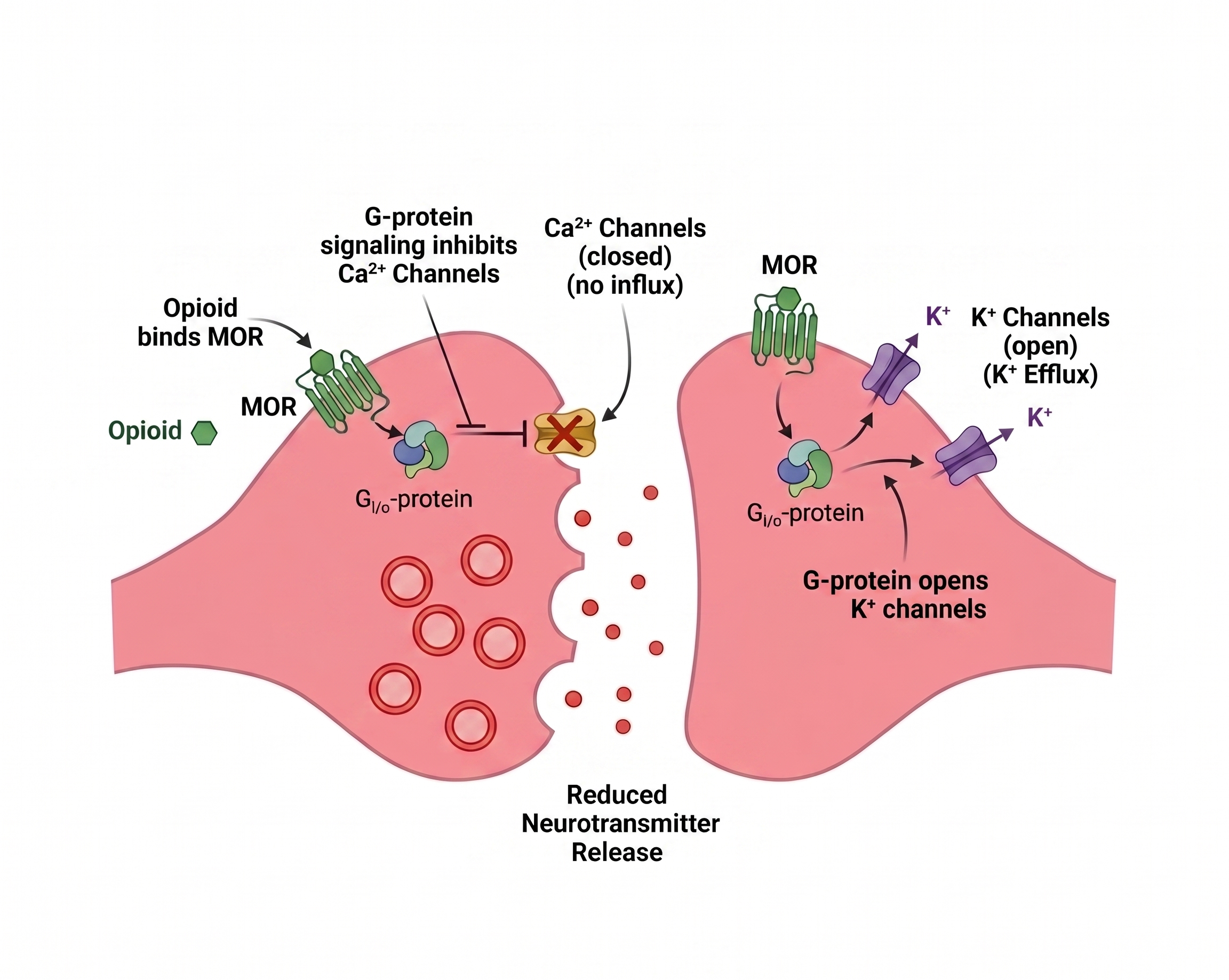
All three are inhibitory G-protein-coupled receptors. They all do the same molecular thing when activated:
- ↓ Intracellular cAMP
- ↑ Potassium channels open - K⁺ flows out.
- ↓ Voltage-gated calcium channels close - Ca²⁺ can't flow in.
Net effect: the neuron is hyperpolarized. The neuron cannot release whatever neurotransmitter it was supposed to release.
Video: Opioids - click to expand ↗ YouTube
Step 2: What MORs Normally Do
MORs evolved to be activated by the brain's own opioids: endorphins, enkephalins, dynorphins. These molecules are released during pain, stress, exercise, social bonding, and orgasm.
Their normal job is to:
- Suppress pain signals in the spinal cord and brainstem.
- Trigger reward in the VTA when survival behaviors succeed.
- Calm breathing in the brainstem when at rest.
Opioid drugs mimic these endorphins.
KOR is activated by dynorphins - the brain's stress/aversion signal. DOR is activated by enkephalins - mood and mild analgesia. Same molecular machinery, different endogenous systems.
Step 3: Where Opioid Receptors Live
MORs are densely expressed in five regions. Each region's normal job is what gets altered.

 and Nucleus Raphe Magnus (Pain img).png)

KOR and DOR live in different places, which is why they produce different effects:


Step 4: How Side Effects Fall Out
For classic MOR agonists (heroin, morphine, fentanyl, oxycodone), each side effect = MOR activation in a specific region altering that region's normal job.
Euphoria
MOR activation in VTA → ↓ GABAergic interneurons (brake released) → ↑ dopamine in NAc. This is disinhibition.
↑
Pain Relief (analgesia)
MOR activation in PAG and spinal cord → ↓ Ca²⁺ at presynaptic terminals → ↓ glutamate & substance P release → pain signal never reaches cortex.
↓
Respiratory Depression - the way opioids kill
MOR activation in preBötzinger Complex → ↓ activity of a small cluster of pacemaker neurons (~70–140 in some estimates; exact count varies across studies - Bachmutsky et al., 2020; Wei & Ramirez, 2019) → rhythm generator goes flat → breathing slows and stops → hypoxia → death. The parabrachial nucleus also contributes meaningfully by providing excitatory drive to the preBötC rhythm generator; suppressing both sites together is how opioids cause respiratory failure (Liu et al., 2021; Varga et al., 2020).
↓
Constipation, Drowsiness, Miosis (pinpoint pupils)
MOR activation in the peripheral nervous system, cortex, and Edinger-Westphal nucleus respectively. Same mechanism, different regions.
Step 5: Why Not All Opioids Are the Same Drug
The framework predicts the difference. Drugs in the same class can feel very different because they engage MOR, KOR, and DOR in different ways.
Flip the MOR switch all the way. No ceiling on euphoria, analgesia, or respiratory depression. These are the deadliest opioids. Fentanyl alone accounted for ~88% of opioid deaths in 2024 (CDC, 2025).
Flips MOR only part of the way no matter the dose. Also blocks KOR, and weakly activates DOR (Volpe et al., 2011). Mechanical consequences:
- Ceiling on respiratory depression - above a certain dose, more drug produces no more breathing suppression because the receptor can't be activated further. Pure buprenorphine overdose is rare.
- High MOR affinity, slow off-rate - sits on MOR a long time and is hard to displace. A person on buprenorphine who takes heroin gets little effect because the receptor is already occupied. This is the basis for using it in addiction treatment.
- KOR antagonism may help with mood during withdrawal by blocking the dysphoria-producing KOR signaling that rises in craving states.
Kratom (Mitragyna speciosa) contains mitragynine, which partially activates MOR. Its active metabolite 7-hydroxymitragynine (7-OH) is a more potent partial MOR agonist than morphine in animal assays (Matsumoto et al., 2004), with meaningful respiratory depression risk at high doses despite being "partial." Kratom is often used for self-managed opioid withdrawal but carries real dependence liability of its own.
Salvinorin A is the cleanest case: a selective high-efficacy KOR agonist (Roth et al., 2002). Same molecular action as MOR drugs, but applied to dopamine terminals. KOR inhibits dopamine release rather than disinhibiting it (Carlezon et al., 2006).
Result: dysphoria, anxiety, depressive-like states instead of euphoria. KOR activation in cortex/claustrum is thought to contribute to dissociative hallucinations. Salvia trips are short (under 15 minutes when smoked) and are usually described as unpleasant, directly traceable to the receptor.
The Balancing Loop
This is MOR-specific, the receptor that drives addiction.
The neuron does not want cAMP suppressed forever. To restore baseline, it manufactures more adenylyl cyclase. A tug-of-war: the drug pushing cAMP ↓, more enzyme pushing it back ↑ (Christie, 2008).

Tolerance. Takes more drug to suppress the elevated enzyme back to "normal." MOR itself also undergoes desensitization, phosphorylation, and internalization that further reduce responsiveness with chronic exposure (Williams et al., 2013).
Withdrawal. Remove the drug → suppression vanishes → upregulated enzyme runs unopposed → cAMP overshoots → neurons in locus coeruleus, raphe magnus, etc. become hyperexcitable → physical symptoms: hyperalgesia, tachycardia, sweating, GI upset, dysphoria. Dynorphin/KOR signaling also rises during withdrawal, adding to the negative emotional state and craving (Koob, 2020).
User Manual
Tolerance to euphoria fades faster than tolerance to respiratory depression, and tolerance to both drops fast during any break (jail, detox, hospital). The dose that produced a high last month can stop breathing today.
After any tolerance break, restart at 25–50% of the previous dose.
Naloxone and xylazine ("tranq"): Naloxone reverses opioid-mediated respiratory depression but has no effect on the alpha-2 adrenergic mechanism of xylazine or medetomidine. If a street opioid contains xylazine, naloxone may restore breathing but wound necrosis and other xylazine-specific harms are not reversed. Give naloxone regardless. It saves lives when opioids are present, but call 911; further medical care may be needed.
Never use alone. Opioid overdose causes unconsciousness before it kills. You cannot self-rescue. If you use alone, tell someone, use a never-use-alone hotline, or have naloxone and an unlocked door where someone can reach you.
"Opioid" is the receptor family, not the experience. A full MOR agonist (heroin) and a KOR agonist (salvia) both fit the definition. They are not the same drug. The question that actually matters: which receptors, how strongly, what kind of agonist.
Sources
- Bachmutsky, I., Wei, X. P., Kish, E., & Yackle, K. (2020). Opioids depress breathing through two small brainstem sites. eLife, 9, e52694. https://doi.org/10.7554/eLife.52694
- Carlezon, W. A., et al. (2006). Depressive-like effects of the κ-opioid receptor agonist salvinorin A on behavior and neurochemistry in rats. Journal of Pharmacology and Experimental Therapeutics, 316(1), 440–447. https://doi.org/10.1124/jpet.105.092304
- Centers for Disease Control and Prevention. (2025). U.S. overdose deaths decrease almost 27% in 2024. National Center for Health Statistics Pressroom. https://www.cdc.gov/nchs/pressroom/releases/20250514.html
- Christie, M. J. (2008). Cellular neuroadaptations to chronic opioids: tolerance, withdrawal and addiction. British Journal of Pharmacology, 154(2), 384–396. https://doi.org/10.1038/bjp.2008.100
- Koob, G. F. (2020). Neurobiology of opioid addiction: opponent process, hyperkatifeia, and negative reinforcement. Biological Psychiatry, 87(1), 44–53. https://doi.org/10.1016/j.biopsych.2019.05.023
- Land, B. B., et al. (2008). The dysphoric component of stress is encoded by activation of the dynorphin κ-opioid system. Journal of Neuroscience, 28(2), 407–414. https://doi.org/10.1523/JNEUROSCI.4458-07.2008
- Matsumoto, K., Horie, S., Ishikawa, H., Takayama, H., Aimi, N., Ponglux, D., & Watanabe, K. (2004). Antinociceptive effect of 7-hydroxymitragynine in mice: Discovery of an orally active opioid analgesic from the Thai medicinal herb Mitragyna speciosa. Life Sciences, 74(17), 2143–2155. https://doi.org/10.1016/j.lfs.2003.09.054
- Neuroscientifically Challenged. (n.d.). 2-Minute neuroscience: Opioids [Video]. YouTube. https://www.youtube.com/watch?v=NPlNCqBHPnE
- Overdose Lifeline. (2024). 2024 opioid overdose data report. https://www.overdoselifeline.org/news/2024-opioid-overdose-data-report-key-trends-and-insights/
- Peppin, J. F., & Raffa, R. B. (2015). Delta opioid agonists: a concise update on potential therapeutic applications. Journal of Clinical Pharmacy and Therapeutics, 40(2), 155–166. https://doi.org/10.1111/jcpt.12244
- Pfeiffer, A., Brantl, V., Herz, A., & Emrich, H. M. (1986). Psychotomimesis mediated by κ opiate receptors. Science, 233(4765), 774–776. https://doi.org/10.1126/science.3016896
- Pradhan, A. A., et al. (2011). The delta opioid receptor: an evolving target for the treatment of brain disorders. Trends in Pharmacological Sciences, 32(10), 581–590. https://doi.org/10.1016/j.tips.2011.06.008
- Roth, B. L., et al. (2002). Salvinorin A: A potent naturally occurring nonnitrogenous κ opioid selective agonist. Proceedings of the National Academy of Sciences, 99(18), 11934–11939. https://doi.org/10.1073/pnas.182234399
- Volpe, D. A., et al. (2011). Uniform assessment and ranking of opioid Mu receptor binding constants for selected opioid drugs. Regulatory Toxicology and Pharmacology, 59(3), 385–390. https://doi.org/10.1016/j.yrtph.2010.12.007
- Liu, Y., et al. (2021). DREADD-mediated activation of subfornical organ neurons suppresses phrenic nerve activity and breathing. eLife, 10, e62503. https://doi.org/10.7554/eLife.62503
- Varga, A. G., et al. (2020). Functional neuroanatomy of the parabrachial nucleus: focus on pneumotaxic mechanisms and opioid-induced respiratory depression. Frontiers in Neuroanatomy, 14, 557003. https://doi.org/10.3389/fnana.2020.557003
- Wei, A. D., & Ramirez, J. M. (2019). Presynaptic mechanisms and KCNQ potassium channels modulate opioid depression of respiratory drive. Frontiers in Physiology, 10, 544. https://doi.org/10.3389/fphys.2019.00544
- Williams, J. T., et al. (2013). Regulation of µ-opioid receptors: desensitization, phosphorylation, internalization, and tolerance. Pharmacological Reviews, 65(1), 223–254. https://doi.org/10.1124/pr.112.005942